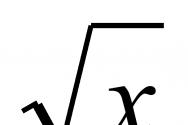Потенциал действия, определения, кривая ПД. Фазы ПД, ионные механизмы их возникновения. Потенциал действия Под потенциалом действия понимают
Потенциалом действия называют быстрое изменение мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых железистых клеток. В основе его возникновения лежат изменения ионной проницаемости мембраны. В развитии потенциала действия выделяют четыре последовательных периода: 1) локальный ответ; 2) деполяризация; 3) реполяризация и 4) следовые потенциалы (рис. 2.11).
Локальный ответ представляет собой активную местную деполяризацию, возникающую вследствие увеличения натриевой проницаемости клеточной мембраны. Уменьшение мембранного потенциала называется деполяризацией. Однако при подпороговом стимуле начальное повышение натриевой проницаемости недостаточно велико, чтобы вызвать быструю деполяризацию мембраны. Локальный ответ возникает не только при подпороговом, но и при надпороговом раз-
Рис. 2.11.
1 - локальный ответ; 2 - фаза деполяризации; 3 - фаза реполяризации; 4 - отрицательный следовой потенциал; 5 - положительный (гиперполяризационный) следовой потенциал
дражении и является составным компонентом потенциала действия. Таким образом, локальный ответ является первоначальной и универсальной формой реагирования ткани на различные по силе раздражения. Биологический смысл локального ответа состоит в том, что если раздражитель по силе мал, то ткань реагирует на него с минимальной тратой энергии, не включая механизмы специфической деятельности. В том же случае, когда раздражение надпороговое, локальный ответ переходит в потенциал действия. Период от начала раздражения до начала фазы деполяризации, когда локальный ответ, нарастая, снижает мембранный потенциал до критического уровня (КУД), называется латентным или скрытым периодом, продолжительность которого зависит от силы раздражения (рис. 2.12).
Фаза деполяризации характеризуется быстрым уменьшением мембранного потенциала и даже перезарядкой мембраны: внутренняя ее часть на некоторое время становится заряженной положительно, а внешняя - отрицательно. Изменение знака заряда на мембране называют извращением - реверсией потенциала. В отличие от локального ответа скорость и величина деполяризации не зависят от силы раздражителя. Продолжительность фазы деполяризации в нервном волокне лягушки составляет около 0,2-0,5 мс.
Продолжительность фазы реполяризации составляет 0,5-0,8 мс. Восстановление исходной величины поляризации мембраны называют реполяризацией. В течение этого времени мембранный потен-

Рис. 2.12. Потенциалы действия, возникающие в ответ на пороговое раздражение коротким (А) и длительным (Б) стимулами Раздражающие стимулы, при воздействии которых получены ответы А и Б: ПП - потенциал покоя; Екуд. - критический уровень деполяризации мембраны (по А.Л. Каталымову)
циал постепенно восстанавливается и достигает 75-85% потенциала покоя. В литературе второй и третий периоды часто называют пиком потенциала действия.
Колебания мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов - следовую деполяризацию и следовую гиперполяризацию, которые соответствуют четвертой и пятой фазам потенциала действия. Следовая деполяризация (отрицательный следовой потенциал) является продолжением фазы реполяризации и характеризуется более медленным (по сравнению с фазой реполяризации) восстановлением потенциала покоя. Следовая деполяризация переходит в следовую гиперполяризацию (положительный следовой потенциал), представляющую собой временное увеличение мембранного потенциала выше исходного уровня. Увеличение мембранного потенциала называется гиперполяризацией. В миелинизирован- ных нервных волокнах следовые потенциалы имеют более сложный характер: следовая деполяризация может переходить в следовую гиперполяризацию, затем иногда возникает новая деполяризация, лишь после этого происходит полное восстановление потенциала покоя.
Ионный механизм возникновения потенциала действия. Основу потенциала действия составляют последовательно развивающиеся во времени изменения ионной проницаемости клеточной мембраны.
При действии на клетку раздражителя проницаемость мембраны для ионов Na + резко повышается за счет активации (открывания) натриевых каналов.
При этом ионы Na + по концентрационному градиенту интенсивно перемещаются из вне во внутриклеточное пространство. Вхождению ионов Na + в клетку способствует и электростатическое взаимодействие. В итоге проницаемость мембраны для Na + становится в 20 раз больше проницаемости для ионов К + .
Сначала деполяризация идет сравнительно медленно. Когда мембранный потенциал уменьшается на 10-40 мВ, скорость деполяризации резко увеличивается и кривая потенциала действия круто поднимается верх. Уровень мембранного потенциала, при котором резко увеличивается скорость деполяризации мембраны, благодаря тому что поток ионов Na + внутрь клетки оказывается большим, чем поток ионов К + наружу, называют критическим уровнем деполяризации.
Поскольку поток Na + в клетку начинает превышать калиевый ток из клетки, происходит постепенное снижение потенциала покоя, приводящее к реверсии - изменению знака мембранного потенциала. При этом внутренняя поверхность мембраны становится электроположительной по отношению к ее внешней электроотрицательной поверхности. Указанные изменения мембранного потенциала соответствуют восходящей фазе потенциала действия (фазе деполяризации).
Мембрана характеризуется повышенной проницаемостью для ионов Na + лишь очень короткое время (0,2-0,5 мс). После этого проницаемость мембраны для ионов Na + вновь понижается, а для К + - возрастает. В результате поток Na + внутрь клетки резко ослабляется, а ток К + из клетки усиливается.
В течение потенциала действия в клетку поступает значительное количество Na + , а ионы К + покидают клетку. Восстановление клеточного ионного баланса осуществляется благодаря работе натрий- калиевого насоса, активность которого возрастает при повышении внутренней концентрации ионов Na + и увеличении внешней концентрации ионов К + . Благодаря работе ионного насоса и изменению проницаемости мембраны для Na + и К + их концентрация во внутри- и внеклеточном пространстве постепенно восстанавливается.
Итогом этих процессов и является реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к внешней поверхности мембраны.
Следовой отрицательный потенциал регистрируется в период, когда № + -каналы инактивированы и реполяризация, связанная с выходом ионов К + из клетки, происходит медленнее, чем во время нисходящей части пика потенциала действия. Это длительное сохранение отрицательности наружной поверхности возбужденного участка по отношению к невозбужденному называют следовой деполяризацией. Следовая деполяризация означает, что в этот период наружная поверхность возбудимого образования имеет меньший положительный заряд, чем в состоянии покоя.
Следовой положительный потенциал соответствует периоду увеличения мембранного потенциала покоя, т.е. гиперполяризации мембраны. Во время следового положительного потенциала наружная поверхность клетки более положительно заряжена, чем в состоянии покоя. Следовой положительный потенциал часто называют следовой гиперполяризацией. Она объясняется длительным сохранением повышенной проницаемости для ионов К + . Вследствие этого на мембране устанавливается потенциал, равный потенциалу равновесия (для К + - 90 мВ).
Изменения возбудимости в процессе развития возбуждения. Воздействуя раздражителями разной силы в различные фазы потенциала действия, можно проследить, как изменяется возбудимость в ходе возбуждения. На рис. 2.13" видно, что период локального ответа характеризуется повышенной возбудимостью (мембранный потенциал приближается к критическому уровню деполяризации); во время фазы деполяризации мембрана утрачивает возбудимость (клетка становится рефрактерной), которая постепенно восстанавливается в ходе реполяризации.
Выделяют период абсолютной рефрактерности , который в нервных клетках продолжается около 1 мс и характеризуется их полной невоз- будимостью. Период абсолютной рефрактерности возникает в результате практически полной инактивации (непроницаемости) натриевых каналов и повышения калиевой проводимости мембраны. Даже в состоянии покоя активированы не все каналы мембраны, 40% из них находятся в состоянии инактивации. При деполяризации количество инактивированных каналов увеличивается и вершина пика потенциала действия соответствует инактивации всех натриевых каналов.
По мере реполяризации мембраны происходит реактивация натриевых каналов. Это период относительной рефрактерности : потенциал действия может возникнуть только при действии более сильных (надпороговых) раздражителей.
В период отрицательного следового потенциала фаза относительной рефрактерности сменяется фазой повышенной (супернормальной) возбудимости. В этот период порог раздражения снижен по сравнению с исходным значением, поскольку мембранный потенциал ближе к критической величине, чем в состоянии покоя (рис. 2.14) .
Фаза следовой гиперполяризации, обусловленная остаточным выходом калия из клетки, напротив, характеризуется снижением

Рис. 2.13.
А - компоненты волны возбуждения: 1 -деполяризация; 2 - реполяризация; МП - мембранный потенциал; мВ - микровольт; МК - критический уровень деполяризации: а - длительность порогового потенциала; б - длительность потенциала действия; в - следовая отрицательность; г - следовая положительность; Б - изменения возбудимости в разные фазы волны возбуждения; УВ - уровень возбудимости в покое: а - повышение возбудимости в период порогового потенциала; б - падение возбудимости до нуля во время протекания потенциала действия (абсолютная рефрактерность); в, - возвращение возбудимости к исходному уровню во время следовой отрицательности (относительная рефрактерность); в 2 - повышение возбудимости в период конца следовой отрицательности (экзальтация"или супернормальность); в - весь период следовой отрицательности; г - падение возбудимости в период гиперполяризации (субнормальность)
возбудимости. Поскольку мембранный потенциал больше, чем в состоянии покоя, требуется более сильный раздражитель для его «смещения» до уровня критической деполяризации.
Таким образом, в динамике возбудительного процесса изменяется способность клетки реагировать на раздражители, т.е. возбудимость.

Рис. 2.14.
Величина мембранного потенциала: Е 0 - в покое; - в фазе экзальтации; Е 2 - в фазе гиперполяризации. Величина порогового потенциала: е 0 - в покое; е, - в фазе экзальтации; е 2 - в фазе гиперполяризации
Это имеет большое значение, поскольку в момент наибольшего возбуждения (пика потенциала действия) клетка становится абсолютно невозбудимой, что защищает ее от гибели и повреждений.
- См.: Леонтьева Н.Н., Маринова К.В. Указ. соч.
- Там же.
Мембранный потенциал (МП) - разность потенциалов между наружной и внутренней стороной мембраны в состоянии физиологического покоя.
Причины возникновения МП:
1. неодинаковое распределение ионов по обе стороны мембраны: внутри - больше К+, снаружи - его мало, но больше Nа+ и Cl. такое распределение ионов называется ионной ассиметрией.
2. избирательная проницаемость мембраны для ионов. В состоянии покоя мембрана неодинакова проницаема.
За счет этих факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта в результате разности концентрации ионов.
Ионы К выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны. Сl - пассивно переходит во внутрь клетки, что приводит к повышению положительного заряда на наружной поверхности мембраны. Nа накапливается на наружной поверхности мембраны и увеличивает «+» заряд. Органические соединения остаются внутри клетки.
В результате такого движения наружная поверхность мембраны «+» заряжена, а внутренняя «-». Внутренняя поверхность может быть «-» заряжена, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние называется поляризацией.
Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов, т.е. пока не наступит электрохимическое равновесие.
Момент равновесия зависит от двух сил:
2. Сила электрохимического взаимодействия.
Значение электрохимического равновесия:
3. поддержание ионной асимметрии
4. поддержание величины мембранного потенциала на постоянном уровне.
Возникновение МП при участи двух сил называют концентрационно-электрохимическим.
Для поддержания ионной симметрии электрохимического равновесия в клетке имеется Nа-К насос. В клеточной мембране имеется система переносчиков, каждый из которых связывает 3Na, которые находятся снаружи, а с внутренней стороны переносчик связывает 2К и переносит внутрь клетки. При этом расходуется 1 молекула АТФ.
Работа Nа-К насоса обеспечивает:
1. высокую концентрацию К внутри клетки, т.е. постоянную величину потенциала покоя
2. низкую концентрацию Nа внутри клетки, т.е. сохраняется нормальная осмомолярность, объем клетки, создает базу для генерации ПД.
3. стабильный концентрационный градиент Nа, способствуя транспорту аминокислот и сахаров.
МП в норме : для гладких мышц -30 - (-70) мВ, для нерва -50 - (-70) мВ, для миокарда -60 - (-90) мВ.
Потенциал действия (ПД) - сдвиг потенциала покоя, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой мембраны.
При действии порогового и сверхпорогового раздражителей изменяется проницаемость клеточной мембраны для ионов. Для Nа увеличивается в 450 раз и градиент нарастает быстро. Для К увеличивается в 10-15 раз и градиент развивается медленно. В результате движение Nа происходит внутрь клетки, К двигается из клетки, что приводит к перезарядке клеточной мембраны.
Фазы:
0. Локальный ответ (местная деполяризация), предшествующий развитию ПД.
1. Фаза деполяризации . Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше 0. Поэтому мембрана приобретает противоположный заряд - внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2 мсек.
2. Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно +20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя. Длительность фазы 3-5 мсек.
3. Фаза следовой деполяризации или следового отрицательного потенциала. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мсек.
4. Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу, МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мсек.
Возникновение ПД обусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые и медленные натриевые каналы. Т.е. натриевая проницаемость мембраны резко возрастает. Причем от чувствительности активационных зависит величина критического уровня деполяризации, чем она выше, тем ниже КУД и наоборот.
Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ), сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия, т.е. калиевый выходящий ток. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации. Возрастание МП приводит к закрыванию и активационных ворот натриевых каналов, что еще больше снижает натриевую проницаемость мембраны и ускоряет реполяризацию.
Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой.
Следовая гиперполяризация связана с повышенной, после ПД, калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос , выносящий вошедшие в клетку во время ПД ионы натрия.
Соотношение фаз потенциала действия и возбудимости.
Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением.
В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности.
В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности.
Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.
В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости. Рис. Следует отметить, что явление аккомодации также связано с изменением проводимости ионных каналов. Если деполяризующий ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит.
Соединение в живых существах органической материи и металлов, использование живой клетки в качестве проводника электрического тока не могут не удивлять. Чем больше подробностей о своем теле я узнаю, тем чаще возникает удивление от простых его действий. Сгибание руки может восприниматься не как привычный, почти не замечаемый процесс, а как результат слаженной работы мышц и сухожилий, получивших команду от мозга, переданную по нервным волокнам с помощью электричества. Разглядывая полосу глубоко переливающейся синевы, возникающей на куске полированного лабрадорита, который я верчу в руках, я воспринимаю излучаемые им электрические волны рецепторами глаз. В рецепторах эта информация кодируется в виде череды электрических импульсов. Она передается в головной мозг, обрабатывается им, а результат всего этого — восприятие таинственно-красивого камня, возникающие при этом эмоции. Матрица, короче:)
Вместе с восхищением возникает и желание узнать — как это все происходит, желание вникать все глубже в подробности каждого процесса.
Потенциал действия — это тот электрический импульс, которой доставляет информацию от рецепторов к мозгу, и с помощью которого мозг управляет телом. Для его существования необходимы особо устроенные клетки, нейроны, мембраны которых облеплены с обеих сторон положительно и отрицательно заряженными ионами, которые только и ждут сигнала, чтобы этот электрический импульс пронести по всей длине клетки. Основным местом существования потенциала действия является аксон нейрона. Дендриты некоторых типов нейронов также способны проводить электрический импульс.

Достаточно взглянуть на картинку слева, чтоб увидеть основное отличие потенциала покоя от потенциала действия: когда мембрана находится в состоянии покоя, на всей ее протяженности снаружи существует положительный электрический заряд, а внутри — отрицательный.

А потенциал действия, проходящий по нейрону от тела клетки до конца аксона, представляет собой изменение внешнего и внутреннего зарядов клеточной мембраны на небольшом участке нейрона: на очень краткий промежуток времени отрицательный и положительный заряды по разным сторонам мембраны меняются местами.
Возникают вопросы: что запускает потенциал действия? Какова его физическая природа, что именно происходит в клетке при его существовании? Каким образом он распространяется? Почему он идет только в одном направлении? Как клетка восстанавливает исходный потенциал покоя после прохождения по ней волны возбуждения?
Итак, первый вопрос: как возникает потенциал действия? Так как передача сигналов от нейронов-источников к нейрону-получателю, или постсинаптическому нейрону (postsynaptic neuron ), происходит в синапсах, запускающий потенциал действия сигнал стоит искать именно там. Через синапс в нейрон приходят сигнальные вещества, но их поступление не обязательно означает запуск возбуждения нейрона. Если бы нейрон реагировал на очень малые количества сигнальных веществ, это было бы так же неудобно, как иметь супер-чувствительный выключатель, срабатывающий, если на него случайно чихнуть.

Необходимо достаточно сильное воздействие определенных сигнальных веществ, чтобы запустить потенциал действия. В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы — временные изменения зарядов мембраны, вызываемые пришедшими через синапс сигнальными веществами .
Постсинаптические потенциалы сильно отличаются от потенциала действия по своей природе. Если потенциал действия имеет одну строго определенную величину, то величина постсинаптических потенциалов зависит только от вида и количества вызвавших их химических веществ, переданных через синапсы. В то время как потенциалы действия, генерируемые разными нейронами, примерно одинаковы, постсинаптические потенциалы, возникающие в разных входных синапсах на одном и том же нейроне, сильно отличаются по величине и продолжительности. В одном синапсе нейрона приходящий нервный импульс может вызвать изменение мембранного потенциала на 0,1 мВ, а в другом — на 20 мВ. Кроме того, существуют как вызывающие потенциал действия, так и тормозящие его сигналы.
Тело клетки служит своеобразным калькулятором, простейшим аналоговым компьютером, суммирующим все поступающие в него сигналы в виде возникающих постсинаптических потенциалов. Аналоговые компьютеры устроены очень просто. Например можно построить водяной аналоговый компьютер. Если из одной трубы в банку вливается три килограмма воды, а из другой в ту же банку — пять, то взвесив банку и выяснив, что весы показывают вес, равный восьми килограммам, мы делаем вывод, что 3+5=8. На заре развития компьютерной техники использовались и водяные, и прочие аналоговые вычислительные машины, но наиболее продвинутой была, конечно, машина, которая в точности как и нейроны суммирует электрические токи. Операторы таких ЭВМ занимались вычислениями, перетыкая штекеры из одного гнезда в другое, сливая вместе разные токи.
Так вот в нейроне происходит такое же суммирование электрических токов. В процессе этого суммирования может быть перейден порог возбуждения (excitation threshold ): будет достигнуто определенное значение мембранного потенциала, которое запускает потенциал действия. Обрати внимание на то, что для достижения порога возбуждения мембранной потенциал должен увеличиваться. Если мембранный потенциал покоя нейрона равен -70 мВ, то его порог возбуждения может быть равен -55 мВ. Снижение мембранного потенциала например до -90 мВ приводит к тому, что для преодоления порога возбуждения требуется более сильное воздействие, ведь он остается прежним: -55мВ.

В постсинаптическом нейроне, получающем сигналы от двух возбуждающих и одного тормозящего нейронов, могут существовать различные комбинации сигналов, и далеко не каждая из них приводит к возникновению потенциала действия.
В первой схеме картинки вверху изображена ситуация, когда только от одного возбуждающего нейрона приходит слабый сигнал, которого недостаточно для преодоления порога возбуждения. Этот сигнал повторяется, но к моменту прихода второго, такого же слабого сигнала от того же аксона возбуждение мембраны уже достигает потенциала покоя, и потенциал действия не возникает.
На второй схеме изображен тот же самый слабый сигнал, но приходящий с большей частотой. Именно из-за увеличения частоты сигнала мембрана не успевает вернуться в состояние покоя в промежутке между двумя сигналами. В этом случае сумма остаточного заряда мембраны после первого сигнала и поступающего следом второго сигнала позволяет преодолеть порог возбуждения нейрона. В этом случае у основания аксона, которое называется аксонным холмиком (axon hillock [‘aksɒn ‘hɪlək] ), возникает потенциал действия.
На третьей схеме от обоих возбуждающих нейронов приходят слабые сигналы, и их суммы достаточно для запуска потенциала действия.
Четвертая схема показывает воздействие на постсинаптический нейрон возбуждающего и тормозящего сигнала. При поступлении только возбуждающего сигнала мембранный потенциал незначительно увеличивается, а при поступлении только тормозного сигнала — снижается. Если эти два сигнала приходят одновременно, то они всего лишь уничтожают друг друга, оставляя мембранный потенциал в состоянии, близком к потенциалу покоя. Это напоминает надавливание на одну и ту же дверь с разных сторон: по отдельности каждое воздействие могло бы приоткрыть дверь или наоборот, прикрыть ее, но существующие одновременно, они приводят лишь к небольшому сдвигу ее по направлению действия большей по модулю силы.
Природа системы такова, что даже малые по величине постсинаптические потенциалы, суммируясь, могут давать большой эффект. Нейрон «складывает» все поступившие в него возбуждающие постсинаптические потенциалы, «вычитает» поступившие тормозные постсинаптические потенциалы, и как только будет перейден порог возбуждения, возникает потенциал действия.
Чтобы разобраться, что происходит в клетке во время запуска и прохождения потенциала действия , необходимо перечислить каждого участвующего игрока и его действие, как было сделано при описании возникновения мембранного потенциала покоя в . Итак, действующие лица при создании потенциала действия :

1 . , которые были закрыты при существовании мембранного потенциала покоя, открываются, как только потенциал достигает порога возбуждения. Так как внутри клетки на мембране существует отрицательный заряд, то в этот момент через каналы туда врываются притягиваемые им
2 . положительно заряженные ионы натрия (Na +). Именно они создавали положительный заряд на внешней поверхности мембраны нейрона, и именно в этот момент они переносят его внутрь, оставляя на внешней мембране отрицательный заряд, который создают находящиеся вне клетки отрицательно заряженные ионы хлора.
3 . таким образом ионы хлора (Cl —) создают отрицательный заряд на внешней поверхности мембраны.

Перемещение ионов натрия внутрь клетки приводит к резкому изменению зарядов внутри и снаружи нейрона. Место существующего во время покоя нейрона отрицательного внутреннего заряда мембраны занимает положительный заряд ионов натрия. А снаружи клеточной мембраны после ухода оттуда Na + наконец-то может проявить себя Cl — , чей отрицательный заряд больше не компенсируется положительно заряженными ионами. Таким образом потенциал клеточной мембраны внезапно возрастает с -70 до +40 мВ. Именно это повышение, называемое деполяризацией (depolarization ), изображено резко идущей вверх частью пика на этой картинке. Такое повышение мембранного потенциала является сигналом закрыть потенциал-зависимые натриевые каналы, а также открывает потенциал-зависимые калиевые каналы, о которых раньше мы еще не говорили.
4 . потенциал-зависимые калиевые каналы. Они существуют в нейроне в дополнение к проточным калиевым каналам, которые, как ты наверное помнишь из прошлой главы про потенциал, не имеют ворот и обеспечивают постоянный отток калия из клетки, играя существенную роль в формировании потенциала покоя. Так вот оказывается, что есть две разновидности калиевых каналов, и сейчас мы рассмотрим работу потенциал-зависимого калиевого канала. Открываясь после того, как потенциал действия достиг своего пика, потенциал-зависимые калиевые каналы резко увеличивают отток ионов калия из клетки. Существующий в этот момент внутри нейрона положительный заряд, образовавшийся за счет притока Na + , активно выпихивает положительно заряженные ионы калия (К +) из клетки через оба вида каналов.
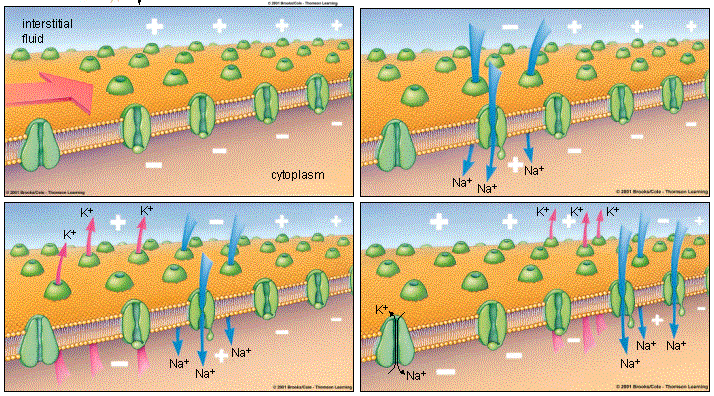
Итак, еще раз проследим цепочку событий до этого момента: потенциал достигает порога возбуждения, и этот электрический сигнал приводит к тому, что открылись потенциал-зависимые натриевые каналы, ионы Na + хлынули внутрь, притягиваемые отрицательно заряженными протеинами, произошла деполяризация, то есть разница зарядов снаружи и внутри резко снизилась, и даже более того — отсутствие положительно заряженных ионов натрия привело к возникновению отрицательного заряда там, где только что был положительный — произошла смена полюсов. Действие электрического импульса закончилось, натриевые каналы вновь закрылись, и столпившиеся внутри ионы Na + вытолкнули одноименно заряженные ионы К + через открывшиеся потенциал-зависимые калиевые каналы и через проточные калиевые каналы.
Заряд внутренней поверхности клеточной мембраны начинает стремительно снижаться, на схеме этому соответствует идущая вниз часть пика. Такое снижение заряда называется реполяризацией (repolarization ), то есть разница зарядов вне и внутри нейрона вновь восстановилась, как и было изначально в состоянии покоя. Но восстановилась именно разница зарядов, а не ионный состав, ее создающий. Несмотря на то, что в конце реполяризации снаружи мембраны образуется положительный заряд, равный положительному заряду мембраны в состоянии покоя, до достижения мембраной состояния покоя еще далеко. Ведь этот положительный заряд принесли туда вышедшие из клетки ионы К + , а при потенциале покоя его поддерживают ионы Na + , которые в данный момент заперты внутри клетки, в том самом месте, где ранее в состоянии покоя находились ионы калия.
В конце реполяризации может возникнуть снижение мембранного потенциала даже ниже его значения для покоящегося нейрона, или гиперполяризация (hyperpolarization [ˌhaɪpəpələraɪ’zeɪʃn] ). Через короткий промежуток времени она компенсируется дополнительным выходом ионов калия из клетки, и наступает прежнее, стабильное состояние покоя.

Когда мембранный потенциал становится равен потенциалу покоя (-70 мВ), потенциал-зависимые калиевые каналы закрываются. Через проточные калиевые каналы К + уже не вытекает так стремительно, ведь его концентрация внутри клетки упала, а снаружи находится много ионов калия. В то же время внутри находится много ионов натрия, которые должны быть выведены из клетки для того, чтоб через нее мог снова пройти потенциал действия.
Восстановлением исходного положения «натрий снаружи, калий внутри» занимается натрий-калиевый насос, выводящий из клетки три Na + и одновременно проносящий внутрь два К + . О его работе было рассказано в предыдущей главе про потенциал, и он схематично показан в четвертой части этой картинки. Совместные действия натрий-калиевого насоса и проточных калиевых каналов приводят к тому, что с внешней стороны мембраны опять возникает большая концентрация Na + , а концентрация К + внутри нейрона выше наружной.
Если бы работали только проточные калиевые каналы, то ионы калия только выходили бы из клетки, и было бы невозможным поддерживать внутри большую концентрацию К + , чем снаружи. Если бы действовали только потенциал-зависимые натриевые каналы, то ионы натрия вообще не могли бы покинуть клетку после того, как вошли туда, создавая потенциал действия. Но натрий-калиевый насос (ну то есть натрий-калиевый канал) решает обе эти проблемы: он выводит наружу Na + , восстанавливая его высокую концентрацию снаружи. И загоняет внутрь прошедшие через проточные каналы К + , не давая им возможности распределиться с обеих сторон мембраны равномерно, лишившись таким образом парциального давления, необходимого для формирования мембранного потенциала покоя.
Через тысячную долю секунды после того, как реполяризация нейронной мембраны достигла минимума, активная работа мембранных каналов восстанавливает потенциал покоя. Нейрон готов к проведению следующего потенциала действия.
5 . положительно заряженные ионы кальция (Ca 2+) играют важную роль в формировании потенциала действия лишь в некоторых нервных клетках, поэтому в данной главе, содержащей общую для различных нейронов информацию, она не будет описана.

На этом общее описание перемещения ионов через клеточную мембрану, превращающее потенциал покоя нейрона в потенциал действия, закончено. Можно сделать перерыв и переварить его вместе с чем-нибудь вкусным, заодно подпитав мозг, компенсировав те калории, которые он затратил на работу по обработке всей этой информации.
Распространение потенциала действия по всей длине аксона обеспечивается тем фактом, что возникший при его существовании в одном участке мембраны нейрона электрический ток возбуждает соседние участки, открывая натриевые потенциал-зависимые каналы и запуская уже там потенциал действия. Точнее, электрический ток запускает потенциал действия только с одной стороны от места существования предыдущего потенциала. Потому что с другой стороны натриевые каналы инактивированы после недавнего открытия, а без их участия возникновение потенциала действия невозможно. Если бы не было этого временного промежутка нефункциональности между двумя открытиями натриевых каналов, потенциал действия не мог бы распространяться только в одном направлении. Происходящая в результате цепная реакция открытия натриевых каналов обеспечивает передачу нервного импульса от места его возникновения до окончаний аксона, подходящих к другим нейронам (или к мышечной клетке).

Схематично эту цепную реакцию можно изобразить так:
Момент времени 1 : Электрический ток, возникший на в результате открытия потенциал-зависимых натриевых каналов на участке 1 , возбуждает соседние участки мембраны.
Момент времени 2 : В результате этого возбуждения на участке 2 открываются потенциал-зависимые натриевые каналы и возникает потенциал действия, который, в свою очередь, возбуждает соседние участки мембраны. На участке 1 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал после прохождения потенциала действия.
Момент времени 3 : Электрический ток, возникший в результате открытия потенциал-зависимых натриевых каналов на участке 2 , вызвал открытие таких же каналов на участке 3 , и потенциал действия перешел туда. На участке 1 потенциал-зависимые натриевые каналы не смогли открыться, несмотря на электрическое возбуждение от участка 2 , так как они были временно инактивированы после создания потенциала действия. На участке 2 открылись потенциал-зависимые калиевые каналы, восстанавливающие мембранный потенциал покоя после прохождения потенциала действия.
Мы уже знаем, что отростки нейронов могут быть покрыты обернутыми вокруг них клетками нейроглии, или миелиновой оболочкой. Именно она делает аксон похожим на вытянутую цепочку сосисок, про ее образование и функции говорилось в . Передача потенциала возбуждения по «голым», не покрытым миелиновыми клетками, нервным волокнам отличается от его прохождения по миелинизированному аксону. При сравнении заметны существенные преимущества, предоставляемые обернутыми вокруг аксона клетками нейроглии:

1 . Экономичность . Мембрана аксона под миелином не имеет каналов для пропускания ионов, создающих потенциал действия. Каналы существуют только в узких промежутках между глиальными клетками, которые называют перехваты Ранвье (Ranvier’s constriction ), и только на этих узких участках аксона возникает потенциал действия. Концентрация ионных каналов в этих перехватах в 100 раз выше, чем в мембранах безмиелиновых волокон, тем не менее для того, чтобы обеспечить их работу, требуется значительно меньше энергии по сравнению с тем, сколько энергии потребовалось бы для обеспечения ионных каналов, расположенных по всей поверхности «голого» аксона. Кроме того, такая экономичность позволяет быстрее «перезаряжать» аксон для проведения следующего потенциала действия, поэтому миелинизированные волокна способны передавать сигналы более высокой частоты.
2
. Скорость
. По «голому» аксону сигнал передается в каждой его точке. Ионные каналы открываются в каждой точке его поверхности, один за другим, что напоминает тщательное, скрупулезное вчитывание в текст.
В миелинезированном аксоне потенциал действия, возникший в одном перехвате Ранвье, своим электрическим полем дотягивается до соседних перехватов, что приводит к возникновению в них новых потенциалов действия. То есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает следующий после поврежденного перехват, а в случае необходимости даже третий, четвертый и пятый по счету. Распространение электрического поля на такую длину возможно из-за электроизоляции, создаваемой миелиновыми муфтами: она уменьшает рассеивание электрического поля, которое неизбежно для безмиелиновых волокон.
Таким образом, скорость распространения потенциала действия по миелинизированным волокнам намного выше по сравнению с немиелинизированными. Ведь за один и тот же отрезок времени потенциал действия на «голом» аксоне возбудит лишь находящийся непосредственно рядом участок, а потенциал действия на миелинизированном волокне перепрыгнет на расстояние, равное расстоянию между одним или несколькими перехватами Ранвье:


Диапазон скоростей передачи потенциала действия в различных миелинизированных волокнах очень широк: от нескольких метров в секунду до «мирового рекорда», установленного аксоном креветки: он проводит возбуждение со скоростью, превышающей 200 м/с.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения потенциала действия за счёт миелиновой оболочки, достаточно сравнить скорость его распространения по немиелинизированным и миелинизированным волокнам нейронов человека. Если взять скорость проведения импульса в одинаково тонких волокнах, то в безмиелиновом волокне она составит примерно 1 м/с, а в миелинизированном 15-20 м/с. В толстых аксонах человека, покрытых миелиновой оболочкой, скорость проведения может достигать 120 м/с.
Итак, подведем итог : передача электрического импульса через тело и отростки нейрона осуществляется с помощью прохождения ионов через клеточную мембрану.

1 . сначала на клеточной мембране создается потенциал покоя . Его образуют положительно заряженные ионы натрия (Na + ) снаружи клетки и отрицательно заряженные протеины внутри клетки. Создающие заряды частицы расположены очень близко к клеточной мембране, ведь они имеют разные знаки и притягиваются друг к другу. Поэтому потенциал покоя существует только возле мембраны, в целом среда внутри нейрона и вне его электрически нейтральна. Кроме Na + и отрицательно заряженных протеинов, в создании и поддержании электрического заряда участвуют положительно заряженные ионы (К + ) и отрицательно заряженные ионы хлора (Cl — ) , тоже находящиеся вблизи мембраны. Потенциал покоя равен примерно -70 мВ (в дальнейшем при указывании численного значения мембранных потенциалов слово «примерно» будет опускаться).
2 . когда нейрон через синапсы получает сигналы от других нейронов, под их влиянием на мембране тела нейрона начинают возникать постсинаптические потенциалы , то есть изменения величины электрических зарядов на внутренней и внешней поверхностях мембраны. Под влиянием этих сигналов внутренний отрицательный заряд и внешний положительный могут увеличиваться или уменьшаться на разные величины. И нейрон выступает в роли калькулятора, суммирующего эти воздействия.

3 . итогом суммирования постсинаптических потенциалов может являться продолжение нахождения нейрона в состоянии покоя. Но если сумма дойдет до значения, называемого порогом возбуждения , то в аксонном холмике (утолщении тела нейрона у основания аксона) возникнет потенциал действия .
4 . в создании потенциала действия главную роль играет клеточная мембрана . Она регулирует прохождение через нее ионов (всех перечисленных, кроме отрицательно заряженных протеинов, которые не могут покидать пределы клетки), открывая и закрывая специализированные каналы, способные проводить только определенные ионы.
5 . когда мембрана нейрона переходит порог возбуждения, в ней открываются потенциал-зависимые натриевые каналы . Исчезает то препятствие, которое отделяло положительно заряженные Na + от притягивающих их отрицательно заряженных протеинов, и Na + устремляются внутрь клетки . Таким образом наружный положительный заряд переносится внутрь клетки, а снаружи остается отрицательный заряд, создаваемый ионами Cl — . Происходит изменение мембранного потенциала от потенциала покоя, равного -70 мВ, до +40 мВ, и это называют потенциалом действия . Важно понимать, что это изменение потенциала происходит только на том участке мембраны, где открылись потенциал-зависимые натриевые каналы.
6 . В момент достижения мембраной потенциала в 40 мВ потенциал-зависимые натриевые каналы закрываются, и на этом же участке открываются потенциал-зависимые калиевые каналы . Снаружи в этот момент существует отрицательный заряд, а внутри — положительный, создаваемый вошедшими туда ионами Na + и уже находящимися там в большом количестве ионами К + . Ионы калия выпихиваются в окружающую нейрон среду через открывшиеся каналы, что резко снижает внутренний заряд мембраны и увеличивает внешний. По обеим сторонам мембраны создаются заряды той же величины, которые обычно поддерживают потенциал покоя. Но это состояние клетки отличается от потенциала покоя тем, что ионы калия и натрия находятся не на своих местах: Na + находится внутри, а К + снаружи. Опять же все описанное касается только одного места на мембране, в котором только что перестал существовать потенциал действия.
7 . Чтобы переместить положительно заряженные ионы Na + и К + по нужные для поддержания потенциала покоя стороны мембраны, используется натрий-калиевый насос . И это действие тоже касается ограниченного участка мембраны: того, где только что закрылись потенциал-зависимые калиевые каналы.
Один цикл работы натрий-калиевого насоса заключается в том, что он захватывает изнутри клетки три иона Na + , перемещает их наружу, где захватывает два иона К + и перемещает их внутрь . То есть канал обменивает три иона Na + на два иона К + , причем Na + могут перемещаться только наружу, а К + только внутрь клетки. Таким образом восстанавливается исходная концентрация натрия на наружной стороне мембраны и калия на внутренней. Натрий-калиевый насос выводит из клетки три положительно заряженных иона, а вводит в нее только два. Следовательно, при каждом его цикле внутренний положительный заряд мембраны уменьшается, а внешний увеличивается. К тому же и вытекающие ионы калия уносят изнутри наружу свои положительные заряды. Возникает вопрос: если потенциал мембраны уже и так был равен потенциалу покоя после входа ионов калия внутрь клетки, зачем его снижать еще сильнее? Ответ заключается в том, что кроме упомянутых здесь ионов, существуют и другие, чье участие в формировании потенциала покоя менее существенно, и именно для компенсации их влияния необходимо такое выравнивание мембранного потенциала. Итогом этого процесса является достижение мембранного потенциала покоя: заряд внутренней стороны мембраны отрицателен, внешней положителен, разность потенциалов равна -70 мВ.
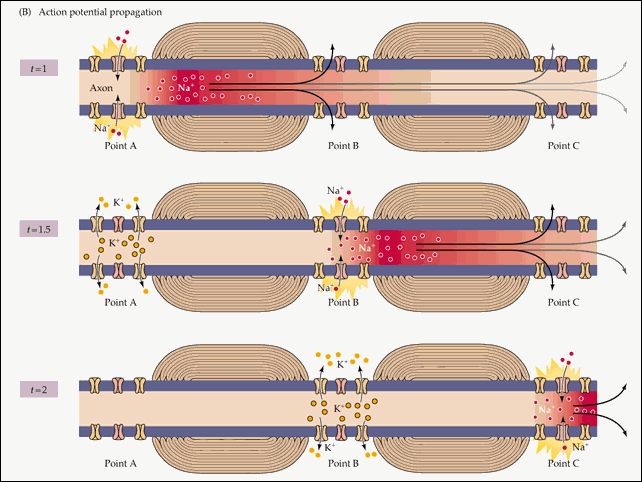
8 . Итак, в том месте клеточной мембраны, где всего три шага назад существовал потенциал действия, опять царит потенциал покоя. Но потенциал действия не прошел бесследно: его существование (описанное в пункте 5) создало электрическое поле, возбудившее соседние с ним части мембраны . С одной стороны (по направлению к концу аксона) это возбуждение вызвало открытие потенциал-зависимых натриевых каналов, запустив уже на новом участке потенциал действия. Который в свою очередь порождает электрическое поле, запускающее следующий потенциал действия, и этот цикл повторяется по всей длине аксона, от его основания до концов, просоединенных к следующим в нервной цепи клеткам.
С другой стороны от потенциала действия, ближе к телу нейрона, несмотря на воздействие электрического поля потенциал действия не возникает. Потому что потенциал-зависимые натриевые каналы на этом участке совсем недавно открывались, и на некоторое время они инактивированы. Именно эта пауза в их работе обеспечивает распространение потенциала действия только в одном направлении.
9 . Любой сигнал, будь то передача информации от сенсорного нейрона в мозг, или команда мозга, идущая к мотонейрону, передается одинаковыми по величине потенциалами действия . Результат потенциала действия (появление восприятия запаха, обусловленное работой сенсорных нейронов, или перемещение тела поближе к его источнику в результате слаженной работы мотонейронов) зависит исключительно от того, в какую нервную цепь включен нейрон, этот потенциал передающий. Для передачи интенсивности сигнала (силы запаха или величины сокращения мышцы) используется частота, с которой потенциалы действия передаются по нейрону. Слабый сигнал передается редкими, а сильный — частыми потенциалами действия .
10 . миелиновая оболочка вокруг отростков нейрона повышает скорость распространения потенциала действия за счет того, что сигнал передается на большое расстояние, равное ширине одной обернутой вокруг отростка клетки нейроглии, а не на непосредственно прилегающий к возбужденному участок, как это происходит у безмиелиновых волокон.
Раздражители
По природе раздражители подразделяют на:
физические (звук, свет, температура, вибрация, осмотическое давление), особое значение для биологических систем имеют электрические раздражители;
химические (ионы, гормоны, нейромедиаторы, пептиды, ксенобиотики);
информационные (голосовые команды, условные знаки, условные стимулы).
По биологическому значению раздражители подразделяют на:
адекватные
– раздражители, для восприятия которых биологическая система имеет специальные приспособления;
неадекватные
– раздражители, не соответствующие природной специализации рецепторных клеток, на которые они действуют.
Раздражитель вызывает возбуждение только в том случае, если он достаточно силен. Порог возбуждения – минимальная сила раздражителя, достаточная для того, чтобы вызвать возбуждение клетки. Выражение «порог возбуждения» имеет несколько синонимов: порог раздражения, пороговая сила раздражителя, порог силы.
Возбуждение как активная реакция клетки на раздражитель
Реакция клетки на внешнее воздействие (раздражение) отличается от реакции небиологических систем следующими особенностями:
энергией для реакции клетки служит не энергия раздражителя, а энергия, образующаяся в результате метаболизма в самой биологической системе;
сила и форма реакции клетки не определяется силой и формой внешнего воздействия (если сила раздражителя выше пороговой).
В некоторых специализированных клетках реакция на раздражитель проявляется особенно интенсивно. Такую интенсивную реакцию называют возбуждением. Возбуждение – активная реакция специализированных (возбудимых) клеток на внешнее воздействие, проявляющаяся в том, что клетка начинает выполнять присущие ей специфические функции.
Возбудимая клетка может находиться в двух дискретных состояниях:
состоянии покоя (готовность к реагированию на внешнее воздействие, совершение внутренней работы);
состоянии возбуждения (активное выполнение специфических функций, совершение внешней работы).
В организме существует 3 типа возбудимых клеток:
нервные клетки (возбуждение проявляется генерацией электрического импульса);
мышечные клетки (возбуждение проявляется сокращением);
секреторные клетки (возбуждение проявляется выбросом в межклеточное пространство биологически активных веществ).
Возбудимость – способность клетки переходить из состояния покоя в состояние возбуждения при действии раздражителя. Разные клетки имеют различную возбудимость. Возбудимость одной и той же клетки меняется в зависимости от ее функционального состояния.
Возбудимая клетка в состоянии покоя
Мембрана возбудимой клетки поляризована. Это означает, что имеется постоянная разность потенциалов между внутренней и наружной поверхностью клеточной мембраны, которую называют мембранный потенциал (МП). В состоянии покоя величина МП составляет –60…–90 мВ (внутренняя сторона мембраны заряжена отрицательно относительно наружной). Значение МП клетки в состоянии покоя называют потенциалом покоя (ПП). МП клетки можно измерять, разместив один электрод внутри, а другой снаружи клетки (рис. 1 А) .
Уменьшение МП относительно его нормального уровня (ПП) называют деполяризацией , а увеличение – гиперполяризацией . Под реполяризацией понимают восстановление исходного уровня МП после его изменения (см. рис. 1 Б).
Электрические и физиологические проявления возбуждения
Рассмотрим различные проявления возбуждения на примере раздражения клетки электрическим током (рис. 2).
При действии слабых (подпороговых) импульсов электрического тока в клетке развивается электротонический потенциал. Электротонический потенциал (ЭП) – сдвиг мембранного потенциала клетки, вызываемый действием постоянного электрического тока . ЭП есть пассивная реакция клетки на электрический раздражитель; состояние ионных каналов и транспорт ионов при этом не изменяется. ЭП не проявляется физиологической реакцией клетки. Поэтому ЭП не является возбуждением.
При действии более сильного подпорогового тока возникает более пролонгированный сдвиг МП – локальный ответ. Локальный ответ (ЛО) – активная реакция клетки на электрический раздражитель, однако состояние ионных каналов и транспорт ионов при этом изменяется незначительно. ЛО не проявляется заметной физиологической реакцией клетки. ЛО называют местным возбуждением , так как это возбуждение не распространяется по мембранам возбудимых клеток.
При действии порогового и сверхпорогового тока в клетке развивается потенциал действия (ПД). ПД характеризуется тем, что значение МП клетки очень быстро уменьшается до 0 (деполяризация), а затем мембранный потенциал приобретает положительное значение (+20…+30 мВ), т. е. внутренняя сторона мембраны заряжается положительно относительно наружной. Затем значение МП быстро возвращается к исходному уровню. Сильная деполяризация клеточной мембраны во время ПД приводит к развитию физиологических проявлений возбуждения (сокращение, секреция и др.). ПД называют распространяющимся возбуждением , поскольку, возникнув в одном участке мембраны, он быстро распространяется во все стороны.
Механизм развития ПД практически одинаков для всех возбудимых клеток. Механизм сопряжения электрических и физиологических проявлений возбуждения различен для разных типов возбудимых клеток (сопряжение возбуждения и сокращения, сопряжение возбуждения и секреции).
Устройство клеточной мембраны возбудимой клетки
В механизмах развития возбуждения участвуют 4 вида ионов: K+ , Na+ , Ca++ , Cl – (ионы Ca++ участвуют в процессах возбуждения некоторых клеток, например кардиомиоцитов, а ионы Cl – важны для развития торможения). Мембрана клетки, представляющая собой липидный бислой, непроницаема для этих ионов. В мембране существуют 2 типа специализированных интегральных белковых систем, которые обеспечивают транспорт ионов через клеточную мембрану: ионные насосы и ионные каналы.
Ионные насосы и трансмембранные ионные градиенты
Ионные насосы (помпы) – интегральные белки, которые обеспечивают активный перенос ионов против градиента концентрации. Энергией для транспорта служит энергия гидролиза АТФ. Различают Na+ / K+ помпу (откачивает из клетки Na+ в обмен на К+), Ca++ помпу (откачивает из клетки Ca++), Cl– помпу (откачивает из клетки Cl –).
В результате работы ионных насосов создаются и поддерживаются трансмембранные ионные градиенты:
концентрация Na+, Ca++, Cl – внутри клетки ниже, чем снаружи (в межклеточной жидкости);
концентрация K+ внутри клетки выше, чем снаружи.
Ионные каналы
Ионные каналы – интегральные белки, которые обеспечивают пассивный транспорт ионов по градиенту концентрации. Энергией для транспорта служит разность концентрации ионов по обе стороны мембраны (трансмембранный ионный градиент).
Неселективные каналы
пропускают все типы ионов, но проницаемость для ионов K+ значительно выше, чем для других ионов;
всегда находятся в открытом состоянии.
Селективные каналы
обладают следующими свойствами:
пропускают только один вид ионов; для каждого вида ионов существует свой вид каналов;
могут находиться в одном из 3 состояний: закрытом, активированном, инактивированном.
Избирательная проницаемость селективного канала обеспечивается селективным фильтром , который образован кольцом из отрицательно заряженных атомов кислорода, которое находится в самом узком месте канала.
Изменение состояния канала обеспечивается работой воротного механизма , который представлен двумя белковыми молекулами. Эти белковые молекулы, так называемые активационные ворота и инактивационные ворота, изменяя свою конформацию, могут перекрывать ионный канал.
В состоянии покоя активационные ворота закрыты, инактивационные ворота открыты (канал закрыт) (рис. 3). При действии на воротную систему сигнала активационные ворота открываются и начинается транспорт ионов через канал (канал активирован). При значительной деполяризации мембраны клетки инактивационные ворота закрываются и транспорт ионов прекращается (канал инактивирован). При восстановлении уровня МП канал возвращается в исходное (закрытое) состояние.
В зависимости от сигнала, который вызывает открытие активационных ворот, селективные ионные каналы подразделяют на:
хемочувствительные каналы
– сигналом к открытию активационных ворот является изменение конформации ассоциированного с каналом белка-рецептора в результате присоединения к нему лиганда;
потенциалчувствительные каналы
– сигналом к открытию активационных ворот является снижение МП (деполяризация) клеточной мембраны до определенного уровня, который называют критическим уровнем деполяризации
(КУД).
Механизм формирования потенциала покоя
Мембранный потенциал покоя образуется главным образом благодаря выходу К+ из клетки через неселективные ионные каналы. Утечка из клетки положительно заряженных ионов приводит к тому, что внутренняя поверхность мембраны клетки заряжается отрицательно относительно наружной.
Мембранный потенциал, возникающий в результате утечки К+ , называют «равновесным калиевым потенциалом» (Ек ). Его можно рассчитать по равнению Нернста

где R
– универсальная газовая постоянная,
Т
– температура (по Кельвину),
F
– число Фарадея,
[К+] нар – концентрация ионов К+ снаружи клетки,
[К+] вн – концентрация ионов К+ внутри клетки.
ПП, как правило, очень близок к Ек, но не точно равен ему. Эта разница объясняется тем, что свой вклад в формирование ПП вносят:
поступление в клетку Na+ и Cl– через неселективные ионные каналы; при этом поступление в клетку Cl– дополнительно гиперполяризует мембрану, а поступление Na+ – дополнительно деполяризует ее; вклад этих ионов в формирование ПП невелик, так как проницаемость неселективных каналов для Cl– и Na + в 2,5 и 25 раза ниже, чем для К+ ;
прямой электрогенный эффект Na+ /К+ ионного насоса, возникающий в том случае, если ионный насос работает асимметрично (количество переносимых в клетку ионов K+ не равно количеству выносимых из клетки ионов Na+).
Механизм развития потенциала действия
В потенциале действия выделяют несколько фаз (рис. 4):
фаза деполяризации;
фаза быстрой реполяризации;
фаза медленной реполяризации (отрицательный следовый потенциал);
фаза гиперполяризации (положительный следовый потенциал).
Фаза деполяризации . Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
Фаза быстрой и медленной реполяризации . В результате деполяризации мембраны происходит открытие потенциалчувствительных К+ -каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется.
Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+ / K+ помпы.
Овершут – период времени, в течение которого мембранный потенциал имеет положительное значение.
Пороговый потенциал – разность между мембранным потенциалом покоя и критическим уровнем деполяризации. Величина порогового потенциала определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
Изменение возбудимости клетки при развитии возбуждения
Если принять уровень возбудимости клетки в состоянии физиологического покоя за норму, то в ходе развития цикла возбуждения можно наблюдать ее колебания. В зависимости от уровня возбудимости выделяют следующие состояния клетки (см. рис. 4).
Супернормальная возбудимость (экзальтация ) – состояние клетки, в котором ее возбудимость выше нормальной. Супернормальная возбудимость наблюдается во время начальной деполяризации и во время фазы медленной реполяризации. Повышение возбудимости клетки в эти фазы ПД обусловлено снижением порогового потенциала по сравнению с нормой.
Абсолютная рефрактерность – состояние клетки, в котором ее возбудимость падает до нуля. Никакой, даже самый сильный, раздражитель не может вызвать дополнительного возбуждения клетки. Во время фазы деполяризации клетка невозбудима, поскольку все ее Na+ -каналы уже находятся в открытом состоянии.
Относительная рефрактерность – состояние, в котором возбудимость клетки значительно ниже нормальной; только очень сильные раздражители могут вызвать возбуждение клетки. Во время фазы реполяризации каналы возвращаются в закрытое состояние и возбудимость клетки постепенно восстанавливается.
Субнормальная возбудимость характеризуется незначительным снижением возбудимости клетки ниже нормального уровня. Это уменьшение возбудимости происходит вследствие возрастания порогового потенциала во время фазы гиперполяризации.
Лекция 2 . Общая физиология возбудимых тканей. Потенциал покоя. Потенциал действия .
۩ Сущность процесса возбуждения . Сущность процесса возбуждения можно сформулировать следующим образом. Все клетки организма имеют электрический заряд, который создается неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различная концентрация анионов и катионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для различных ионов и работы ионных насосов. Процесс возбуждения начинается с действия на возбудимую клетку раздражителя. Сначала очень быстро повышается проницаемость её мембраны для ионов натрия и быстро возвращается к норме, затем – для ионов калия и также быстро, но с некоторым отставанием возвращается к норме. Вследствие этого ионы перемещаются в клетку и из клетки согласно электрохимическому градиенту – это и есть процесс возбуждения. Возбуждение возможно только в том случае, если клетка постоянно поддерживает потенциал покоя (мембранный потенциал) и при её раздражении быстро изменяется проницаемость клеточной мембраны.
۩ Потенциал покоя . Потенциал покоя (ПП) – это разность электрических потенциалов между внутренней и наружной средами клетки в состоянии её покоя. При этом внутри клетки регистрируется отрицательный заряд. Величина ПП в различных клетках различна. Так, в волокнах скелетной мышцы регистрируется ПП равный 60-90 мВ, в нейронах – 50-80 мВ, в гладких мышцах – 30-70 мВ, в сердечной мышце – 80-90 мВ. Органеллы клеток имеют свои вариабельные мембранные потенциалы.
Непосредственной причиной существования потенциала покоя является неодинаковая концентрация анионов и катионов внутри и вне клетки (смотри таб.1!).
Таблица 1. Внутри- и внеклеточные концентрации ионов в мышечных клетках.
|
Внутриклеточная концентрация, мМ |
Внеклеточная концентрация, мМ |
|||
|
A- (крупномолекулярные внутриклеточные анионы) |
A-(крупномолекулярные внутриклеточные анионы) |
|||
|
Небольшое количество |
Небольшое количество |
|||
|
Очень мало |
Основное количество |
|||
Неравномерное расположение ионов внутри и вне клетки является следствием неодинаковой проницаемости клеточной мембраны для различных ионов и работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки электрохимическому градиенту. Проницаемость – это её способность пропускать воду, незаряженные и заряженные частицы согласно законам диффузии и фильтрации. Она определяется:
Размерами каналов и размерами частиц;
Растворимостью частиц в мембране (клеточная мембрана проницаема для растворимых в ней липидов и непроницаема для пептидов).
Проводимость – это способность заряженных частиц проходить через клеточную мембрану согласно электрохимическому градиенту.
Различная проницаемость различных ионов играет важную роль в формировании ПП:
Калий является основным ионом, обеспечивающим формирование ПП, так как его проницаемость в 100 раз выше, чем проницаемость для натрия. При уменьшении концентрации калия в клетке ПП уменьшается, а при увеличении – увеличивается. Он может входить и выходить из клетки. В покое количество входящих ионов калия и выходящих его ионов уравновешивается и устанавливается так называемый калиевый равновесный потенциал, который рассчитывается по уравнению Нернста. Механизм его таков: так как электрический и конценрационный градиенты противодействуют друг другу, то калий стремится выйти наружу по концентрационному градиенту, а отрицательный заряд внутри клетки и положительный вне клетки препятствует этому. Тогда количество входящих ионов становится равным количеству выходящих ионов.
Натрий входит в клетку. Его проницаемость мала по сравнению с проницаемостью калия, поэтому его вклад в формирование ПП невелик.
Хлор входит в клетку в незначительном количестве, так как проницаемость мембраны для него невелика, причем он уравновешивается количеством ионов натрия (противоположные заряды притягиваются). Следовательно, его вклад в формирование ПП невелик.
Органические анионы (глютамат, аспартат, органические фосфаты, сульфаты) вообще не могут выйти из клетки, так как они имеют большие размеры. Поэтому за счет них внутри клетки формируется отрицательный заряд.
Роль ионов кальция в формировании ПП заключается в том, что они взаимодействуют с наружными отрицательными зарядами мембраны клетки и отрицательными карбоксильными группами интерстиция, нейтрализуя их, что ведет к стабилизации ПП.
Кроме выше перечисленных ионов, в формировании ПП играют важную роль и поверхностные заряды мембраны (в основном отрицательные). Их формируют гликопротеиды, гликолипиды и фосфолипиды: фиксированные наружные отрицательные заряды, нейтрализуя положительные заряды внешней поверхности мембраны, уменьшают ПП, а фиксированные внутренние отрицательные заряды мембраны, напротив, увеличивают ПП, суммируясь с анионами внутри клетки. Таким образом, потенциал покоя - это алгебраическая сумма всех положительных и отрицательных зарядов ионов вне и внутри клетки и поверхностных зарядов клеточной мембраны .
Роль ионных насосов в формировании ПП . Ионный насос – это молекула белка, которая обеспечивает перенос иона с непосредственной затратой энергии вопреки электрическому и концентрационному градиентам. В результате сопряженного транспорта натрия и калия поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ обеспечивает один цикл работы Na/K-насоса – перенос трех ионов натрия за пределы клетки и двух ионов калия внутрь клетки. Таким образом, увеличивается ПП. Нормальная величина потенциала покоя является необходимым условием для формирования потенциала действия, то есть для формирования процесса возбуждения.
۩Потенциал действия . Потенциал действия – это электрофизиологический процесс, который выражается в быстром колебании мембранного потенциала вследствие изменения проницаемости мембраны и диффузии ионов в клетку и из клетки. Роль ПД заключается в обеспечении передачи сигналов между нервными клетками, нервными центрами и рабочими органами, в мышцах ПД обеспечивает процесс электромеханического сопряжения. ПД подчиняется закону «всё или ничего». Если сила раздражения мала, то возникает локальный потенциал, который не распространяется.
Потенциал действия состоит из трех фаз: деполяризации, то есть исчезновения ПП; инверсии – изменения знака заряда клетки на обратный; реполяризации – восстановление исходного МП.
Механизм возникновения потенциала действия .
Фаза деполяризации . При действии раздражителя на клетку начальная частичная деполяризация клеточной мембраны происходит без изменения ее проницаемости для ионов. Когда деполяризация достигает примерно 50% пороговой величины, возрастает проницаемость мембраны для Na + , причем в первый момент сравнительно медленно. В этот период движущей силой, обеспечивающей движение Na + в клетку, являются концентрационный и электрический градиенты. Вспомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются), а концентрация Na + вне клетки в 12 раз больше, чем внутри клетки. Условием, обеспечивающим дальнейший вход Na + в клетку, является увеличение проницаемости клеточной мембраны, который определяется состоянием воротного механизма натриевых каналов. Воротный механизм натриевых каналов расположен на внешней и внутренней стороне клеточной мембраны, воротный механизм калиевых каналов – только на внутренней стороне мембраны. В каналах для натрия имеются активационные m-ворота, которые расположены с внешней стороны клеточной мембраны, и инактивационные h-ворота, расположенные с внутренней стороны мембраны. В условиях покоя активационные m-ворота закрыты, инактивационные h-ворота открыты. Калиевые активационные ворота закрыты, а инактивационных калиевых ворот нет. Когда деполяризация клетки достигает критической величины, которая обычно составляет 50 мВ, проницаемость мембраны для Na + резко возрастает, так как открывается большое количество потенциалзависимых m-ворот натриевых каналов и ионы натрия лавиной устремляются в клетку. Развивающаяся деполяризация клеточной мембраны вызывает дополнительное увеличение ее проницаемости и, соответственно, проводимости натрия: открываются все новые и новые активационные m-ворота. В итоге ПП исчезает, то есть становится равным нулю. Фаза деполяризации на этом заканчивается. Ее длительность составляет примерно 0,2-0,5 мс.
Фаза инверсии . Процесс перезарядки мембраны представляет собой вторую фазу ПД – фазу инверсии. Фаза инверсии делится на восходящую и нисходящую составляющие. Восходящая часть . После исчезновения ПП вход в клетку ионов натрия продолжается, так как натриевые активационные m-ворота еще открыты. В результате заряд внутри клетки становится положительным, а снаружи-отрицательным. В течение доли миллисекунды ионы натрия еще продолжают входить в клетку. Таким образом, вся восходящая часть пика ПД обеспечивается в основном входом Na + в клетку. Нисходящая составляющая фазы инверсии . Примерно через 0,2-0,5 мс после начала деполяризации рост ПД прекращается в результате закрытия натриевых инактивационных h-ворот и открытия калиевых активационных ворот. Поскольку калий находится преимущественно внутри клетки, он, согласно концентрационному градиенту, начинает быстро выходить из нее, вследствие чего уменьшается число положительно заряженных ионов в клетке. Заряд клетки снова начинает уменьшаться. Во время нисходящей составляющей фазы инверсии выходу ионов калия из клетки способствует также и электрический градиент. К + выталкивается положительным зарядом из клетки и притягивается отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки. Калий выходит из клетки не только по управляемым каналам, но и по неуправляемым каналам – каналам утечки. Амплитуда ПД складывается из величины ПП и величины фазы инверсии, составляющей у разных клеток 10-50 мВ.
Фаза реполяризации . Пока активационные калиевые каналы открыты, K + еще продолжает выходить из клетки, согласно химическому градиенту. Заряд внутри клетки становится отрицательным, а снаружи – положительным, следовательно, электрический градиент резко тормозит выход ионов калия из клетки. Но так как сила химического градиента больше силы электрического градиента, ионы калия продолжают очень медленно выходить из клетки. Затем активационные калиевые ворота закрываются, остается только выход ионов калия по каналам утечки, то есть по концентрационному градиенту через неуправляемые каналы.
Таким образом, ПД вызывается циклическим процессом поступления ионов натрия в клетку и последующего выхода калия из нее. Роль Са 2+ в возникновении ПД в нервных клетках незначительна. Однако Са 2+ играет очень важную роль в возникновении ПД сердечной мышцы, в передаче импульсов от одного нейрона к другому, от нервного волокна к мышечному, в обеспечении мышечного сокращения.
Вслед за ПД возникают следовые явления (характерные для нейронов) – сначала следовая гиперполяризация, а затем следовая деполяризация. Следовая гиперполяризация клеточной мембраны обычно является следствием еще сохраняющейся повышенной проницаемости мембраны для ионов калия. Следовая деполяризация связана с кратковременным повышением проницаемости мембраны для Na + и входом его в клетку согласно химическому и электрическому градиентам.
Кроме этого существуют: а) так называемая фаза абсолютной рефрактерности , или полная невозбудимость клетки. Она приходится на пик ПД и продолжатся 1-2 мс; и б) фаза относительной рефрактерности – период частичного восстановления клетки, когда сильное раздражение может вызвать новое возбуждение. Относительная рефрактерность соответствует конечной части фазы реполяризации и следовой гиперполяризации клеточной мембраны. В нейронах вслед за гиперполяризацией возможна частичная деполяризация клеточной мембраны. В этот период очередной потенциал действия можно вызвать более слабым раздражением, так как МП несколько меньше обычного. Этот период называется фазой экзальтации (период повышенной возбудимости).
Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность. Лабильность , или функциональная подвижность, - это скорость протекания одного цикла возбуждения. Мерой лабильности возбудимого образования является максимальное число ПД, которое он может воспроизвести в 1 секунду. Обычно возбуждение продолжается менее 1 мс и подобно взрыву. Такой «взрыв» протекает мощно, но быстро завершается.
Мышцы, отличияДокумент
... . Возбудимость ткани и ее мера. Законы раздражения возбудимых тканей : силы, времени действия раздражителя... потенциал покоя (МПП); 2) мембранный потенциал действия (МПД); 3) потенциал градиента основного обмена (метаболический потенциал ). Потенциал ...